Making PUFAs Available for Tissue Protection – Selective Phospholipase-Mediated Membrane Release of Omega-3 LCPUFA
This article at a glance 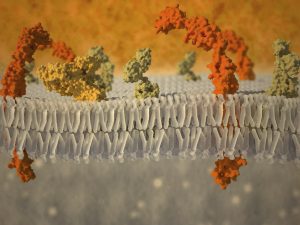 One phospholipase enzyme, the cytosolic phospholipase A2 (cPLA2 or group IVA PLA2), has long been regarded as an important phospholipase that releases arachidonic acid (AA) from membranes. This occurs after a cell is stimulated (e.g. by exposure to a specific hormone, or upon recognition of a microbe in the case of an immune cell). After cPLA2 activation, the free fatty acids formed are further metabolized by different oxygenases that “couple” with phospholipase-mediated release to form fatty acid metabolites, also called lipid mediators, each with specific biological activities. The precise sum of lipid mediators that is formed by this combination of phospholipase and biosynthetic enzymes depends on which enzymes are expressed in the vicinity of the site of PUFA release. Stimulus-coupled AA-release for eicosanoid (AA-derived lipid mediators) generation generally refers to an intracellular event. The lipid mediators that are produced are frequently exported/secreted and exert potent extracellular modulatory and signalling functions in the tissue where they are produced. They act as locally acting hormones (autacoids) with very short half-lives that modulate the function of cells within the organ where they are formed, carrying information on cellular status. For example, an immune cell may sense an infectious bacterium, triggering phospholipase-mediated release of AA, which is transformed into an eicosanoid that activates another cell to act against the bacterium by secreting an anti-microbial peptide. In addition to intracellular phospholipases, several secretory phospholipases secreted from cells have extracellular actions. One of these is group X sPLA2 (sPLA2-X/sPLA2G10). sPLA2-X hydrolyzes AA from zwitterionic membrane phospholipids. Studies have revealed that sPLA2-X has anti-viral and anti-bacterial activity, can hydrolyze platelet-activating factor (an important pro-inflammatory mediator), and also releases AA from intracellular and extracellular membrane sites for the formation of eicosanoids. Several studies have implicated a role for sPLA2-X in different inflammatory settings.
One phospholipase enzyme, the cytosolic phospholipase A2 (cPLA2 or group IVA PLA2), has long been regarded as an important phospholipase that releases arachidonic acid (AA) from membranes. This occurs after a cell is stimulated (e.g. by exposure to a specific hormone, or upon recognition of a microbe in the case of an immune cell). After cPLA2 activation, the free fatty acids formed are further metabolized by different oxygenases that “couple” with phospholipase-mediated release to form fatty acid metabolites, also called lipid mediators, each with specific biological activities. The precise sum of lipid mediators that is formed by this combination of phospholipase and biosynthetic enzymes depends on which enzymes are expressed in the vicinity of the site of PUFA release. Stimulus-coupled AA-release for eicosanoid (AA-derived lipid mediators) generation generally refers to an intracellular event. The lipid mediators that are produced are frequently exported/secreted and exert potent extracellular modulatory and signalling functions in the tissue where they are produced. They act as locally acting hormones (autacoids) with very short half-lives that modulate the function of cells within the organ where they are formed, carrying information on cellular status. For example, an immune cell may sense an infectious bacterium, triggering phospholipase-mediated release of AA, which is transformed into an eicosanoid that activates another cell to act against the bacterium by secreting an anti-microbial peptide. In addition to intracellular phospholipases, several secretory phospholipases secreted from cells have extracellular actions. One of these is group X sPLA2 (sPLA2-X/sPLA2G10). sPLA2-X hydrolyzes AA from zwitterionic membrane phospholipids. Studies have revealed that sPLA2-X has anti-viral and anti-bacterial activity, can hydrolyze platelet-activating factor (an important pro-inflammatory mediator), and also releases AA from intracellular and extracellular membrane sites for the formation of eicosanoids. Several studies have implicated a role for sPLA2-X in different inflammatory settings.  It is recognized that selectivity in the release of different PUFA types by specific phospholipases functions in combination with stimulus-dependent coupling of such release with particular lipid mediator biosynthetic enzymes. Additionally, some phospholipases have in recent years been recognized to selectively release omega-3 long-chain polyunsaturated fatty acids (LCPUFA) such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) to enable their transformation into specific bioactive lipid mediators. The group IID secretory PLA2 (sPLA2G2D) was documented not long ago to selectively release AA for the formation of a specific prostaglandin D2-derived anti-inflammatory metabolite (15-deoxy-Δ12,14-prostaglandin J2), alongside the release of DHA to favor the formation of the inflammation-resolving autacoid resolvin D1. These lipid mediators were found to contribute to protection against experimental contact dermatitis and the control of dendritic cell activity. In a similar fashion in murine skin, group IIF PLA2 (PLA2G2F) preferentially releases DHA to drive the formation of protectin D1, another DHA-derived pro-resolving lipid mediator. A recent study has now uncovered a new role for the group X sPLA2 in the release of omega-3 LCPUFA coupled to the formation of lipid mediators that serve an anti-inflammatory role in the colon of mice. The study by Murase and colleagues was carried out at the Tokyo Metropolitan Institute of Medical Science, with colleagues at the Department of Pharmacy at Showa University, and various other institutions in Tokyo, Japan. The implications of this recognition are likely to be profound since the results suggest that membrane pools of specific PUFA are accessible selectively to specific phospholipase enzymes as important points of control towards regulating specific cellular functions. First, the researchers carried out a detailed characterization of the immune function and anatomy of a mouse that carried a transgenic construct of the human group X sPLA2. These mice overexpressing the enzyme had a smaller heart, and a smaller spleen with reduced white pulp areas (sites of lymphocyte development), containing fewer B-cells with a different stage of differentiation than splenic B-cells in normal wild type mice. Serum levels of the IgG1 and IgE antibody types were increased. T-lymphocytes were less mature than in wild type mice, and the levels of T-lymphocytes in the circulation were markedly reduced. In the thymus, several genes that are involved in lymphocyte maturation, proliferation and migration were expressed at lower levels. Generalized overexpression of transgenically-expressed human group X sPLA2 in mice thus led to an altered immune phenotype.
It is recognized that selectivity in the release of different PUFA types by specific phospholipases functions in combination with stimulus-dependent coupling of such release with particular lipid mediator biosynthetic enzymes. Additionally, some phospholipases have in recent years been recognized to selectively release omega-3 long-chain polyunsaturated fatty acids (LCPUFA) such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) to enable their transformation into specific bioactive lipid mediators. The group IID secretory PLA2 (sPLA2G2D) was documented not long ago to selectively release AA for the formation of a specific prostaglandin D2-derived anti-inflammatory metabolite (15-deoxy-Δ12,14-prostaglandin J2), alongside the release of DHA to favor the formation of the inflammation-resolving autacoid resolvin D1. These lipid mediators were found to contribute to protection against experimental contact dermatitis and the control of dendritic cell activity. In a similar fashion in murine skin, group IIF PLA2 (PLA2G2F) preferentially releases DHA to drive the formation of protectin D1, another DHA-derived pro-resolving lipid mediator. A recent study has now uncovered a new role for the group X sPLA2 in the release of omega-3 LCPUFA coupled to the formation of lipid mediators that serve an anti-inflammatory role in the colon of mice. The study by Murase and colleagues was carried out at the Tokyo Metropolitan Institute of Medical Science, with colleagues at the Department of Pharmacy at Showa University, and various other institutions in Tokyo, Japan. The implications of this recognition are likely to be profound since the results suggest that membrane pools of specific PUFA are accessible selectively to specific phospholipase enzymes as important points of control towards regulating specific cellular functions. First, the researchers carried out a detailed characterization of the immune function and anatomy of a mouse that carried a transgenic construct of the human group X sPLA2. These mice overexpressing the enzyme had a smaller heart, and a smaller spleen with reduced white pulp areas (sites of lymphocyte development), containing fewer B-cells with a different stage of differentiation than splenic B-cells in normal wild type mice. Serum levels of the IgG1 and IgE antibody types were increased. T-lymphocytes were less mature than in wild type mice, and the levels of T-lymphocytes in the circulation were markedly reduced. In the thymus, several genes that are involved in lymphocyte maturation, proliferation and migration were expressed at lower levels. Generalized overexpression of transgenically-expressed human group X sPLA2 in mice thus led to an altered immune phenotype.  Interestingly, the immune status of resident macrophages in the abdominal cavity was found to be highly polarized to an anti-inflammatory phenotype. When the mice were stimulated to mount an acute inflammatory response, a marked inhibition of inflammation was observed (approximately 75% reduction in macrophage infiltration). Furthermore, macrophages isolated from the bone-marrow displayed an anti-inflammatory phenotype upon stimulation. In addition to the lymphocyte immunosuppression and the anti-inflammatory polarization of macrophages, the mice were found to be leaner than wild type mice, and had reduced visceral and subcutaneous adipose tissue. In order to understand how overexpression of human group X sPLA2 could confer such an anti-inflammatory phenotype to mice, the investigators measured a range of lipid mediators derived from the PUFA that sPLA2 might be releasing from cellular membranes. In the spleen, the enhanced release of AA, EPA and DHA was confirmed. Only small non-significant increases in AA-derived eicosanoids were found, but the formation of several EPA- and DHA-derived mediators was much greater than in control mice. In the skin, there was increased release of DHA and formation of DHA-derived protectin D1. And in colonic tissue, there was no change in the formation of AA-derived lipid mediators, but a significant increase in EPA-derived 12-HEPE. Taken together, the results in this mouse model suggested that human group X sPLA2 is able to mobilize omega-3 LCPUFA, in addition to and possibly more efficiently than AA, and appeared to direct these PUFA towards the formation of various anti-inflammatory autacoids. The researchers next turned to studying the role of the endogenous murine group X phospholipase A2 homologous enzyme, sPLA2-X, in wild type mice and in mice lacking this enzyme. Several types of phospholipase A2s were found to be naturally expressed in mouse colon, among which sPLA2-X. After inducing experimental colitis (by introduction of a strong irritant in the drinking water), a more marked reduction in body weight over a 1-week period was found in mice that lacked sPLA2-X compared to wild type mice. Diarrhea and blood present in feces was significantly worse in mice with colitis when the enzyme was absent. The protective effect of sPLA2-X was also seen upon histological examination with reduced epithelial damage, ulceration and submucosal inflammation in mice that had normal sPLA2-X expression compared to the knock-out mice. Deficiencies in any of the other phospholipase enzymes normally found in the colon did not confer this susceptible phenotype in the colitis model. Mice that lacked sPLA2-X and were challenged by experimental colitis displayed a marked upregulation of several cytokines, chemokines and macrophage markers, all associated with a marked pro-inflammatory reaction. Protection from colitis was subsequently shown to be due to contributions from sPLA2-X expressed in colonic epithelium. An additional contribution from sPLA2-X present in hematopoietic cells furthermore appeared to contribute to tissue protection, particularly for other effects such as a splenic response to injury and the maintenance of hematocrit. Analysis of endogenously produced lipid mediators during colitis showed that mice with normal sPLA2-X expression displayed production of higher levels of several EPA- and DHA-derived lipid mediators with pro-resolving activity (18-HEPE, resolvin D2, 4-HDHA and 7-HDHA) than knock-out mice. The formation of several prostaglandins, thromboxane and 12-HHT, all AA-derived lipid mediators, was however unaltered in mice lacking sPLA2-X. The release of EPA, docosapentaenoic acid (DPA) omega-3 and DHA was significantly higher in wild type mice with colitis than in the sPLA2-X knock-out mice, but the release of AA was increased to a much smaller extent. Next, the authors tested if the administration of various PUFA and several select lipid mediators could modify the inflammatory responsiveness of lymphocytes isolated from colonic tissue from wild type mice with colitis. All tested PUFA (AA, EPA, DPA and DHA) reduced the secretion of pro-inflammatory cytokines. The anti-inflammatory effect on lymphocytes was stimulated by activation of the GPR120 receptor, but was not activated by some of the downstream lipid mediators generated from EPA or DHA (at least not by the resolvins that were tested). Daily rectal installation of EPA and DHA reduced inflammation and restored body weight loss in mice lacking sPLA2-X with colitis. Taken together, the results suggest that sPLA2-X may play an important role in mediating the release of omega-3 LCPUFA to mediate anti-inflammatory and tissue protective actions. On lymphocytes these anti-inflammatory actions may be mediated directly by the free fatty acids. The authors did consider that some of the detected omega-3 LCPUFA-derived lipid mediators may be exerting tissue-protective and anti-inflammatory actions on immune cell types other than lamina propria lymphocytes, but this was not tested.
Interestingly, the immune status of resident macrophages in the abdominal cavity was found to be highly polarized to an anti-inflammatory phenotype. When the mice were stimulated to mount an acute inflammatory response, a marked inhibition of inflammation was observed (approximately 75% reduction in macrophage infiltration). Furthermore, macrophages isolated from the bone-marrow displayed an anti-inflammatory phenotype upon stimulation. In addition to the lymphocyte immunosuppression and the anti-inflammatory polarization of macrophages, the mice were found to be leaner than wild type mice, and had reduced visceral and subcutaneous adipose tissue. In order to understand how overexpression of human group X sPLA2 could confer such an anti-inflammatory phenotype to mice, the investigators measured a range of lipid mediators derived from the PUFA that sPLA2 might be releasing from cellular membranes. In the spleen, the enhanced release of AA, EPA and DHA was confirmed. Only small non-significant increases in AA-derived eicosanoids were found, but the formation of several EPA- and DHA-derived mediators was much greater than in control mice. In the skin, there was increased release of DHA and formation of DHA-derived protectin D1. And in colonic tissue, there was no change in the formation of AA-derived lipid mediators, but a significant increase in EPA-derived 12-HEPE. Taken together, the results in this mouse model suggested that human group X sPLA2 is able to mobilize omega-3 LCPUFA, in addition to and possibly more efficiently than AA, and appeared to direct these PUFA towards the formation of various anti-inflammatory autacoids. The researchers next turned to studying the role of the endogenous murine group X phospholipase A2 homologous enzyme, sPLA2-X, in wild type mice and in mice lacking this enzyme. Several types of phospholipase A2s were found to be naturally expressed in mouse colon, among which sPLA2-X. After inducing experimental colitis (by introduction of a strong irritant in the drinking water), a more marked reduction in body weight over a 1-week period was found in mice that lacked sPLA2-X compared to wild type mice. Diarrhea and blood present in feces was significantly worse in mice with colitis when the enzyme was absent. The protective effect of sPLA2-X was also seen upon histological examination with reduced epithelial damage, ulceration and submucosal inflammation in mice that had normal sPLA2-X expression compared to the knock-out mice. Deficiencies in any of the other phospholipase enzymes normally found in the colon did not confer this susceptible phenotype in the colitis model. Mice that lacked sPLA2-X and were challenged by experimental colitis displayed a marked upregulation of several cytokines, chemokines and macrophage markers, all associated with a marked pro-inflammatory reaction. Protection from colitis was subsequently shown to be due to contributions from sPLA2-X expressed in colonic epithelium. An additional contribution from sPLA2-X present in hematopoietic cells furthermore appeared to contribute to tissue protection, particularly for other effects such as a splenic response to injury and the maintenance of hematocrit. Analysis of endogenously produced lipid mediators during colitis showed that mice with normal sPLA2-X expression displayed production of higher levels of several EPA- and DHA-derived lipid mediators with pro-resolving activity (18-HEPE, resolvin D2, 4-HDHA and 7-HDHA) than knock-out mice. The formation of several prostaglandins, thromboxane and 12-HHT, all AA-derived lipid mediators, was however unaltered in mice lacking sPLA2-X. The release of EPA, docosapentaenoic acid (DPA) omega-3 and DHA was significantly higher in wild type mice with colitis than in the sPLA2-X knock-out mice, but the release of AA was increased to a much smaller extent. Next, the authors tested if the administration of various PUFA and several select lipid mediators could modify the inflammatory responsiveness of lymphocytes isolated from colonic tissue from wild type mice with colitis. All tested PUFA (AA, EPA, DPA and DHA) reduced the secretion of pro-inflammatory cytokines. The anti-inflammatory effect on lymphocytes was stimulated by activation of the GPR120 receptor, but was not activated by some of the downstream lipid mediators generated from EPA or DHA (at least not by the resolvins that were tested). Daily rectal installation of EPA and DHA reduced inflammation and restored body weight loss in mice lacking sPLA2-X with colitis. Taken together, the results suggest that sPLA2-X may play an important role in mediating the release of omega-3 LCPUFA to mediate anti-inflammatory and tissue protective actions. On lymphocytes these anti-inflammatory actions may be mediated directly by the free fatty acids. The authors did consider that some of the detected omega-3 LCPUFA-derived lipid mediators may be exerting tissue-protective and anti-inflammatory actions on immune cell types other than lamina propria lymphocytes, but this was not tested.  The study addresses some additional points, including a demonstration that selective release of AA by cPLA2 and subsequent formation of prostaglandin E2 (PGE2) occurs in parallel to the uncovered action of sPLA2-X, and contributes to the tissue protective response towards colitis. The overall conclusion of the study is that cPLA2 and sPLA2-X exert a protective effect against colitis by facilitating the formation of distinct lipid metabolites derived from AA and from omega-3 LCPUFA (EPA, DPA and DHA), respectively. In addition to the colon, sPLA2-X was found to be expressed in the sperm cell acrosome, where the enzyme selectively liberated DPA omega-3 and DHA from the sperm cell membrane after capacitation (activation of sperm cells permitting ovum fertilization). This study offers a fundamental insight into the role of specific phospholipases in being able to selectively release specific PUFA species and couple this release to important downstream functions, such as shown here in anti-inflammation, tissue protection, and sperm cell fertility. This study supports the notion that substrate specificity (fatty acid selectivity) of a phospholipase is possibly even more important than the absolute and relative content of EPA/DHA versus AA bound within membrane phospholipid pools, on the premise that sufficient PUFA substrate is available, to appropriately couple the release to cell-type specific downstream lipid mediator formation and action. In previous studies sPLA2-X was shown to play an aggravating role and also an inhibitory role in the development of atherosclerosis. It now remains to be determined if such divergent outcomes depend on the relative availability of EPA/DHA versus AA in the membranes on which sPLA2-X acts. An anti-inflammatory role for the human sPLA2-X in macrophages had been previously reported, and although release of omega-3 LCPUFA-derived lipid mediators was not determined in that study, the enzyme was able to couple AA release to the formation of AA-derived lipid mediators with anti-inflammatory activity. Although further research is needed to demonstrate the importance of the relative availability of EPA/DHA versus AA availability, this observation may mean that the same enzyme is able to also use AA and couple this release to specific AA-derived lipid mediators that also mediate an anti-inflammatory activity.
The study addresses some additional points, including a demonstration that selective release of AA by cPLA2 and subsequent formation of prostaglandin E2 (PGE2) occurs in parallel to the uncovered action of sPLA2-X, and contributes to the tissue protective response towards colitis. The overall conclusion of the study is that cPLA2 and sPLA2-X exert a protective effect against colitis by facilitating the formation of distinct lipid metabolites derived from AA and from omega-3 LCPUFA (EPA, DPA and DHA), respectively. In addition to the colon, sPLA2-X was found to be expressed in the sperm cell acrosome, where the enzyme selectively liberated DPA omega-3 and DHA from the sperm cell membrane after capacitation (activation of sperm cells permitting ovum fertilization). This study offers a fundamental insight into the role of specific phospholipases in being able to selectively release specific PUFA species and couple this release to important downstream functions, such as shown here in anti-inflammation, tissue protection, and sperm cell fertility. This study supports the notion that substrate specificity (fatty acid selectivity) of a phospholipase is possibly even more important than the absolute and relative content of EPA/DHA versus AA bound within membrane phospholipid pools, on the premise that sufficient PUFA substrate is available, to appropriately couple the release to cell-type specific downstream lipid mediator formation and action. In previous studies sPLA2-X was shown to play an aggravating role and also an inhibitory role in the development of atherosclerosis. It now remains to be determined if such divergent outcomes depend on the relative availability of EPA/DHA versus AA in the membranes on which sPLA2-X acts. An anti-inflammatory role for the human sPLA2-X in macrophages had been previously reported, and although release of omega-3 LCPUFA-derived lipid mediators was not determined in that study, the enzyme was able to couple AA release to the formation of AA-derived lipid mediators with anti-inflammatory activity. Although further research is needed to demonstrate the importance of the relative availability of EPA/DHA versus AA availability, this observation may mean that the same enzyme is able to also use AA and couple this release to specific AA-derived lipid mediators that also mediate an anti-inflammatory activity.  This emerging appreciation of selectivity in the release of PUFA by distinct phospholipases in combination with downstream stimulus-dependent coupling towards specific lipid mediator profiles provides us with a reminder that PUFA biology is highly regulated and complex. Making careful assessments of the influence of both omega-3 and omega-6 status of a cell or tissue, the involvement of distinct phospholipases, and the panel of mediators formed and that activate specific cellular reactions, will over time provide a better understanding of the programs that our biological system employs to regulate defined processes in physiology. Murase R, Sato H, Yamamoto K, Ushida A, Nishito Y, Ikeda K, Kobayashi T, Yamamoto T, Taketomi Y, Murakami M. Group X secreted phospholipase A2 releases ω3 polyunsaturated fatty acids, suppresses colitis, and promotes sperm fertility. J. Biol. Chem. 2016;291(13):6895-6911. [PubMed] Worth Noting Ait-Oufella H, Herbin O, Lahoute C, Coatrieux C, Loyer X, et al. Group X secreted phospholipase A2 limits the development of atherosclerosis in LDL receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 2013;33(3):466-473. [PubMed] Curfs DM, Ghesquiere SA, Vergouwe MN, van der Made I, Gijbels MJ, Greaves DR, Verbeek JS, Hofker MH, de Winther MP. Macrophage secretory phospholipase A2 group X enhances anti-inflammatory responses, promotes lipid accumulation, and contributes to aberrant lung pathology. J. Biol. Chem. 2008;283(31):21640-21648. [PubMed] Dennis EA, Cao J, Hsu YH, Magrioti V, Kokotos G. Phospholipase A2 enzymes: physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011;111(10):6130-6185. [PubMed] Jemel I, Ii H, Oslund RC, Payre C, Dabert-Gay AS, Douguet D, Chargui K, Scarzello S, Gelb MH, Lambeau G. Group X secreted phospholipase A2 proenzyme is matured by a furin-like proprotein convertase and releases arachidonic acid inside of human HEK293 cells. J. Biol. Chem. 2011;286(42):36509-36521. [PubMed] Miki Y, Yamamoto K, Taketomi Y, Sato H, Shimo K, Kobayashi T, Ishikawa Y, Ishii T, Nakanishi H, Ikeda K, Taguchi R, Kabashima K, Arita M, Arai H, Lambeau G, Bollinger JM, Hara S, Gelb MH, Murakami M. Lymphoid tissue phospholipase A2 group IID resolves contact hypersensitivity by driving antiinflammatory lipid mediators. J. Exp. Med. 2013;210(6):1217-1234. [PubMed] Mouchlis VD, Bucher D, McCammon JA, Dennis EA. Membranes serve as allosteric activators of phospholipase A2, enabling it to extract, bind, and hydrolyze phospholipid substrates. Proc. Natl. Acad. Sci. U.S.A. 2015;112(6):E516-525. [PubMed] Pruzanski W, Lambeau L, Lazdunsky M, Cho W, Kopilov J, Kuksis A. Differential hydrolysis of molecular species of lipoprotein phosphatidylcholine by groups IIA, V and X secretory phospholipases A2. Biochim. Biophys. Acta 2005;1736(1):38-50. [PubMed] Saiga A, Uozumi N, Ono T, Seno K, Ishimoto Y, Arita H, Shimizu T, Hanasaki K. Group X secretory phospholipase A2 can induce arachidonic acid release and eicosanoid production without activation of cytosolic phospholipase A2 alpha. Prostaglandins Other Lipid Mediat. 2005;75(1-4):79-89. [PubMed] Tremblay BL, Cormier H, Rudkowska I, Lemieux S, Couture P, Vohl MC. Association between polymorphisms in phospholipase A2 genes and the plasma triglyceride response to an n-3 PUFA supplementation: a clinical trial. Lipids Health Dis. 2015;14:12. [PubMed] Tremblay BL, Rudkowska I, Couture P, Lemieux S, Julien P, Vohl MC. Modulation of C-reactive protein and plasma omega-6 fatty acid levels by phospholipase A2 gene polymorphisms following a 6-week supplementation with fish oil. Prostaglandins Leukot. Essent. Fatty Acids 2015;102-103:37-45. [PubMed] Yamamoto K, Miki Y, Sato M, Taketomi Y, Nishito Y, Taya C, Muramatsu K, Ikeda K, Nakanishi H, Taguchi R, Kambe N, Kabashima K, Lambeau G, Gelb MH, Murakami M. The role of group IIF-secreted phospholipase A2 in epidermal homeostasis and hyperplasia. J. Exp. Med. 2015;212(11):1901-1919. [PubMed]
This emerging appreciation of selectivity in the release of PUFA by distinct phospholipases in combination with downstream stimulus-dependent coupling towards specific lipid mediator profiles provides us with a reminder that PUFA biology is highly regulated and complex. Making careful assessments of the influence of both omega-3 and omega-6 status of a cell or tissue, the involvement of distinct phospholipases, and the panel of mediators formed and that activate specific cellular reactions, will over time provide a better understanding of the programs that our biological system employs to regulate defined processes in physiology. Murase R, Sato H, Yamamoto K, Ushida A, Nishito Y, Ikeda K, Kobayashi T, Yamamoto T, Taketomi Y, Murakami M. Group X secreted phospholipase A2 releases ω3 polyunsaturated fatty acids, suppresses colitis, and promotes sperm fertility. J. Biol. Chem. 2016;291(13):6895-6911. [PubMed] Worth Noting Ait-Oufella H, Herbin O, Lahoute C, Coatrieux C, Loyer X, et al. Group X secreted phospholipase A2 limits the development of atherosclerosis in LDL receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 2013;33(3):466-473. [PubMed] Curfs DM, Ghesquiere SA, Vergouwe MN, van der Made I, Gijbels MJ, Greaves DR, Verbeek JS, Hofker MH, de Winther MP. Macrophage secretory phospholipase A2 group X enhances anti-inflammatory responses, promotes lipid accumulation, and contributes to aberrant lung pathology. J. Biol. Chem. 2008;283(31):21640-21648. [PubMed] Dennis EA, Cao J, Hsu YH, Magrioti V, Kokotos G. Phospholipase A2 enzymes: physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011;111(10):6130-6185. [PubMed] Jemel I, Ii H, Oslund RC, Payre C, Dabert-Gay AS, Douguet D, Chargui K, Scarzello S, Gelb MH, Lambeau G. Group X secreted phospholipase A2 proenzyme is matured by a furin-like proprotein convertase and releases arachidonic acid inside of human HEK293 cells. J. Biol. Chem. 2011;286(42):36509-36521. [PubMed] Miki Y, Yamamoto K, Taketomi Y, Sato H, Shimo K, Kobayashi T, Ishikawa Y, Ishii T, Nakanishi H, Ikeda K, Taguchi R, Kabashima K, Arita M, Arai H, Lambeau G, Bollinger JM, Hara S, Gelb MH, Murakami M. Lymphoid tissue phospholipase A2 group IID resolves contact hypersensitivity by driving antiinflammatory lipid mediators. J. Exp. Med. 2013;210(6):1217-1234. [PubMed] Mouchlis VD, Bucher D, McCammon JA, Dennis EA. Membranes serve as allosteric activators of phospholipase A2, enabling it to extract, bind, and hydrolyze phospholipid substrates. Proc. Natl. Acad. Sci. U.S.A. 2015;112(6):E516-525. [PubMed] Pruzanski W, Lambeau L, Lazdunsky M, Cho W, Kopilov J, Kuksis A. Differential hydrolysis of molecular species of lipoprotein phosphatidylcholine by groups IIA, V and X secretory phospholipases A2. Biochim. Biophys. Acta 2005;1736(1):38-50. [PubMed] Saiga A, Uozumi N, Ono T, Seno K, Ishimoto Y, Arita H, Shimizu T, Hanasaki K. Group X secretory phospholipase A2 can induce arachidonic acid release and eicosanoid production without activation of cytosolic phospholipase A2 alpha. Prostaglandins Other Lipid Mediat. 2005;75(1-4):79-89. [PubMed] Tremblay BL, Cormier H, Rudkowska I, Lemieux S, Couture P, Vohl MC. Association between polymorphisms in phospholipase A2 genes and the plasma triglyceride response to an n-3 PUFA supplementation: a clinical trial. Lipids Health Dis. 2015;14:12. [PubMed] Tremblay BL, Rudkowska I, Couture P, Lemieux S, Julien P, Vohl MC. Modulation of C-reactive protein and plasma omega-6 fatty acid levels by phospholipase A2 gene polymorphisms following a 6-week supplementation with fish oil. Prostaglandins Leukot. Essent. Fatty Acids 2015;102-103:37-45. [PubMed] Yamamoto K, Miki Y, Sato M, Taketomi Y, Nishito Y, Taya C, Muramatsu K, Ikeda K, Nakanishi H, Taguchi R, Kambe N, Kabashima K, Lambeau G, Gelb MH, Murakami M. The role of group IIF-secreted phospholipase A2 in epidermal homeostasis and hyperplasia. J. Exp. Med. 2015;212(11):1901-1919. [PubMed]
- A recent study in mice reports that human and mouse secretory group X phospholipase A2 preferentially releases omega-3 long-chain polyunsaturated fatty acids to serve anti-inflammatory and tissue-protective activities in the colon.
- Selective release of arachidonic acid by a different phospholipase, coupled to the biosynthesis of prostaglandin E2, occurs in parallel, and also contributes to the tissue-protective activity.
- Growing recognition of the involvement of distinct phospholipases in specific immunological events may one day allow us to better support their activities to drive beneficial outcomes.
One phospholipase enzyme, the cytosolic phospholipase A2 (cPLA2 or group IVA PLA2), has long been regarded as an important phospholipase that releases arachidonic acid (AA) from membranes. This occurs after a cell is stimulated (e.g. by exposure to a specific hormone, or upon recognition of a microbe in the case of an immune cell). After cPLA2 activation, the free fatty acids formed are further metabolized by different oxygenases that “couple” with phospholipase-mediated release to form fatty acid metabolites, also called lipid mediators, each with specific biological activities. The precise sum of lipid mediators that is formed by this combination of phospholipase and biosynthetic enzymes depends on which enzymes are expressed in the vicinity of the site of PUFA release. Stimulus-coupled AA-release for eicosanoid (AA-derived lipid mediators) generation generally refers to an intracellular event. The lipid mediators that are produced are frequently exported/secreted and exert potent extracellular modulatory and signalling functions in the tissue where they are produced. They act as locally acting hormones (autacoids) with very short half-lives that modulate the function of cells within the organ where they are formed, carrying information on cellular status. For example, an immune cell may sense an infectious bacterium, triggering phospholipase-mediated release of AA, which is transformed into an eicosanoid that activates another cell to act against the bacterium by secreting an anti-microbial peptide. In addition to intracellular phospholipases, several secretory phospholipases secreted from cells have extracellular actions. One of these is group X sPLA2 (sPLA2-X/sPLA2G10). sPLA2-X hydrolyzes AA from zwitterionic membrane phospholipids. Studies have revealed that sPLA2-X has anti-viral and anti-bacterial activity, can hydrolyze platelet-activating factor (an important pro-inflammatory mediator), and also releases AA from intracellular and extracellular membrane sites for the formation of eicosanoids. Several studies have implicated a role for sPLA2-X in different inflammatory settings. It is recognized that selectivity in the release of different PUFA types by specific phospholipases functions in combination with stimulus-dependent coupling of such release with particular lipid mediator biosynthetic enzymes. Additionally, some phospholipases have in recent years been recognized to selectively release omega-3 long-chain polyunsaturated fatty acids (LCPUFA) such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) to enable their transformation into specific bioactive lipid mediators. The group IID secretory PLA2 (sPLA2G2D) was documented not long ago to selectively release AA for the formation of a specific prostaglandin D2-derived anti-inflammatory metabolite (15-deoxy-Δ12,14-prostaglandin J2), alongside the release of DHA to favor the formation of the inflammation-resolving autacoid resolvin D1. These lipid mediators were found to contribute to protection against experimental contact dermatitis and the control of dendritic cell activity. In a similar fashion in murine skin, group IIF PLA2 (PLA2G2F) preferentially releases DHA to drive the formation of protectin D1, another DHA-derived pro-resolving lipid mediator. A recent study has now uncovered a new role for the group X sPLA2 in the release of omega-3 LCPUFA coupled to the formation of lipid mediators that serve an anti-inflammatory role in the colon of mice. The study by Murase and colleagues was carried out at the Tokyo Metropolitan Institute of Medical Science, with colleagues at the Department of Pharmacy at Showa University, and various other institutions in Tokyo, Japan. The implications of this recognition are likely to be profound since the results suggest that membrane pools of specific PUFA are accessible selectively to specific phospholipase enzymes as important points of control towards regulating specific cellular functions. First, the researchers carried out a detailed characterization of the immune function and anatomy of a mouse that carried a transgenic construct of the human group X sPLA2. These mice overexpressing the enzyme had a smaller heart, and a smaller spleen with reduced white pulp areas (sites of lymphocyte development), containing fewer B-cells with a different stage of differentiation than splenic B-cells in normal wild type mice. Serum levels of the IgG1 and IgE antibody types were increased. T-lymphocytes were less mature than in wild type mice, and the levels of T-lymphocytes in the circulation were markedly reduced. In the thymus, several genes that are involved in lymphocyte maturation, proliferation and migration were expressed at lower levels. Generalized overexpression of transgenically-expressed human group X sPLA2 in mice thus led to an altered immune phenotype. Interestingly, the immune status of resident macrophages in the abdominal cavity was found to be highly polarized to an anti-inflammatory phenotype. When the mice were stimulated to mount an acute inflammatory response, a marked inhibition of inflammation was observed (approximately 75% reduction in macrophage infiltration). Furthermore, macrophages isolated from the bone-marrow displayed an anti-inflammatory phenotype upon stimulation. In addition to the lymphocyte immunosuppression and the anti-inflammatory polarization of macrophages, the mice were found to be leaner than wild type mice, and had reduced visceral and subcutaneous adipose tissue. In order to understand how overexpression of human group X sPLA2 could confer such an anti-inflammatory phenotype to mice, the investigators measured a range of lipid mediators derived from the PUFA that sPLA2 might be releasing from cellular membranes. In the spleen, the enhanced release of AA, EPA and DHA was confirmed. Only small non-significant increases in AA-derived eicosanoids were found, but the formation of several EPA- and DHA-derived mediators was much greater than in control mice. In the skin, there was increased release of DHA and formation of DHA-derived protectin D1. And in colonic tissue, there was no change in the formation of AA-derived lipid mediators, but a significant increase in EPA-derived 12-HEPE. Taken together, the results in this mouse model suggested that human group X sPLA2 is able to mobilize omega-3 LCPUFA, in addition to and possibly more efficiently than AA, and appeared to direct these PUFA towards the formation of various anti-inflammatory autacoids. The researchers next turned to studying the role of the endogenous murine group X phospholipase A2 homologous enzyme, sPLA2-X, in wild type mice and in mice lacking this enzyme. Several types of phospholipase A2s were found to be naturally expressed in mouse colon, among which sPLA2-X. After inducing experimental colitis (by introduction of a strong irritant in the drinking water), a more marked reduction in body weight over a 1-week period was found in mice that lacked sPLA2-X compared to wild type mice. Diarrhea and blood present in feces was significantly worse in mice with colitis when the enzyme was absent. The protective effect of sPLA2-X was also seen upon histological examination with reduced epithelial damage, ulceration and submucosal inflammation in mice that had normal sPLA2-X expression compared to the knock-out mice. Deficiencies in any of the other phospholipase enzymes normally found in the colon did not confer this susceptible phenotype in the colitis model. Mice that lacked sPLA2-X and were challenged by experimental colitis displayed a marked upregulation of several cytokines, chemokines and macrophage markers, all associated with a marked pro-inflammatory reaction. Protection from colitis was subsequently shown to be due to contributions from sPLA2-X expressed in colonic epithelium. An additional contribution from sPLA2-X present in hematopoietic cells furthermore appeared to contribute to tissue protection, particularly for other effects such as a splenic response to injury and the maintenance of hematocrit. Analysis of endogenously produced lipid mediators during colitis showed that mice with normal sPLA2-X expression displayed production of higher levels of several EPA- and DHA-derived lipid mediators with pro-resolving activity (18-HEPE, resolvin D2, 4-HDHA and 7-HDHA) than knock-out mice. The formation of several prostaglandins, thromboxane and 12-HHT, all AA-derived lipid mediators, was however unaltered in mice lacking sPLA2-X. The release of EPA, docosapentaenoic acid (DPA) omega-3 and DHA was significantly higher in wild type mice with colitis than in the sPLA2-X knock-out mice, but the release of AA was increased to a much smaller extent. Next, the authors tested if the administration of various PUFA and several select lipid mediators could modify the inflammatory responsiveness of lymphocytes isolated from colonic tissue from wild type mice with colitis. All tested PUFA (AA, EPA, DPA and DHA) reduced the secretion of pro-inflammatory cytokines. The anti-inflammatory effect on lymphocytes was stimulated by activation of the GPR120 receptor, but was not activated by some of the downstream lipid mediators generated from EPA or DHA (at least not by the resolvins that were tested). Daily rectal installation of EPA and DHA reduced inflammation and restored body weight loss in mice lacking sPLA2-X with colitis. Taken together, the results suggest that sPLA2-X may play an important role in mediating the release of omega-3 LCPUFA to mediate anti-inflammatory and tissue protective actions. On lymphocytes these anti-inflammatory actions may be mediated directly by the free fatty acids. The authors did consider that some of the detected omega-3 LCPUFA-derived lipid mediators may be exerting tissue-protective and anti-inflammatory actions on immune cell types other than lamina propria lymphocytes, but this was not tested. The study addresses some additional points, including a demonstration that selective release of AA by cPLA2 and subsequent formation of prostaglandin E2 (PGE2) occurs in parallel to the uncovered action of sPLA2-X, and contributes to the tissue protective response towards colitis. The overall conclusion of the study is that cPLA2 and sPLA2-X exert a protective effect against colitis by facilitating the formation of distinct lipid metabolites derived from AA and from omega-3 LCPUFA (EPA, DPA and DHA), respectively. In addition to the colon, sPLA2-X was found to be expressed in the sperm cell acrosome, where the enzyme selectively liberated DPA omega-3 and DHA from the sperm cell membrane after capacitation (activation of sperm cells permitting ovum fertilization). This study offers a fundamental insight into the role of specific phospholipases in being able to selectively release specific PUFA species and couple this release to important downstream functions, such as shown here in anti-inflammation, tissue protection, and sperm cell fertility. This study supports the notion that substrate specificity (fatty acid selectivity) of a phospholipase is possibly even more important than the absolute and relative content of EPA/DHA versus AA bound within membrane phospholipid pools, on the premise that sufficient PUFA substrate is available, to appropriately couple the release to cell-type specific downstream lipid mediator formation and action. In previous studies sPLA2-X was shown to play an aggravating role and also an inhibitory role in the development of atherosclerosis. It now remains to be determined if such divergent outcomes depend on the relative availability of EPA/DHA versus AA in the membranes on which sPLA2-X acts. An anti-inflammatory role for the human sPLA2-X in macrophages had been previously reported, and although release of omega-3 LCPUFA-derived lipid mediators was not determined in that study, the enzyme was able to couple AA release to the formation of AA-derived lipid mediators with anti-inflammatory activity. Although further research is needed to demonstrate the importance of the relative availability of EPA/DHA versus AA availability, this observation may mean that the same enzyme is able to also use AA and couple this release to specific AA-derived lipid mediators that also mediate an anti-inflammatory activity. This emerging appreciation of selectivity in the release of PUFA by distinct phospholipases in combination with downstream stimulus-dependent coupling towards specific lipid mediator profiles provides us with a reminder that PUFA biology is highly regulated and complex. Making careful assessments of the influence of both omega-3 and omega-6 status of a cell or tissue, the involvement of distinct phospholipases, and the panel of mediators formed and that activate specific cellular reactions, will over time provide a better understanding of the programs that our biological system employs to regulate defined processes in physiology. Murase R, Sato H, Yamamoto K, Ushida A, Nishito Y, Ikeda K, Kobayashi T, Yamamoto T, Taketomi Y, Murakami M. Group X secreted phospholipase A2 releases ω3 polyunsaturated fatty acids, suppresses colitis, and promotes sperm fertility. J. Biol. Chem. 2016;291(13):6895-6911. [PubMed] Worth Noting Ait-Oufella H, Herbin O, Lahoute C, Coatrieux C, Loyer X, et al. Group X secreted phospholipase A2 limits the development of atherosclerosis in LDL receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 2013;33(3):466-473. [PubMed] Curfs DM, Ghesquiere SA, Vergouwe MN, van der Made I, Gijbels MJ, Greaves DR, Verbeek JS, Hofker MH, de Winther MP. Macrophage secretory phospholipase A2 group X enhances anti-inflammatory responses, promotes lipid accumulation, and contributes to aberrant lung pathology. J. Biol. Chem. 2008;283(31):21640-21648. [PubMed] Dennis EA, Cao J, Hsu YH, Magrioti V, Kokotos G. Phospholipase A2 enzymes: physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011;111(10):6130-6185. [PubMed] Jemel I, Ii H, Oslund RC, Payre C, Dabert-Gay AS, Douguet D, Chargui K, Scarzello S, Gelb MH, Lambeau G. Group X secreted phospholipase A2 proenzyme is matured by a furin-like proprotein convertase and releases arachidonic acid inside of human HEK293 cells. J. Biol. Chem. 2011;286(42):36509-36521. [PubMed] Miki Y, Yamamoto K, Taketomi Y, Sato H, Shimo K, Kobayashi T, Ishikawa Y, Ishii T, Nakanishi H, Ikeda K, Taguchi R, Kabashima K, Arita M, Arai H, Lambeau G, Bollinger JM, Hara S, Gelb MH, Murakami M. Lymphoid tissue phospholipase A2 group IID resolves contact hypersensitivity by driving antiinflammatory lipid mediators. J. Exp. Med. 2013;210(6):1217-1234. [PubMed] Mouchlis VD, Bucher D, McCammon JA, Dennis EA. Membranes serve as allosteric activators of phospholipase A2, enabling it to extract, bind, and hydrolyze phospholipid substrates. Proc. Natl. Acad. Sci. U.S.A. 2015;112(6):E516-525. [PubMed] Pruzanski W, Lambeau L, Lazdunsky M, Cho W, Kopilov J, Kuksis A. Differential hydrolysis of molecular species of lipoprotein phosphatidylcholine by groups IIA, V and X secretory phospholipases A2. Biochim. Biophys. Acta 2005;1736(1):38-50. [PubMed] Saiga A, Uozumi N, Ono T, Seno K, Ishimoto Y, Arita H, Shimizu T, Hanasaki K. Group X secretory phospholipase A2 can induce arachidonic acid release and eicosanoid production without activation of cytosolic phospholipase A2 alpha. Prostaglandins Other Lipid Mediat. 2005;75(1-4):79-89. [PubMed] Tremblay BL, Cormier H, Rudkowska I, Lemieux S, Couture P, Vohl MC. Association between polymorphisms in phospholipase A2 genes and the plasma triglyceride response to an n-3 PUFA supplementation: a clinical trial. Lipids Health Dis. 2015;14:12. [PubMed] Tremblay BL, Rudkowska I, Couture P, Lemieux S, Julien P, Vohl MC. Modulation of C-reactive protein and plasma omega-6 fatty acid levels by phospholipase A2 gene polymorphisms following a 6-week supplementation with fish oil. Prostaglandins Leukot. Essent. Fatty Acids 2015;102-103:37-45. [PubMed] Yamamoto K, Miki Y, Sato M, Taketomi Y, Nishito Y, Taya C, Muramatsu K, Ikeda K, Nakanishi H, Taguchi R, Kambe N, Kabashima K, Lambeau G, Gelb MH, Murakami M. The role of group IIF-secreted phospholipase A2 in epidermal homeostasis and hyperplasia. J. Exp. Med. 2015;212(11):1901-1919. [PubMed]